The main focus of considerable scientific research has been to understand exactly how cell function changes with age to deduce how cells age. Many theories of aging have been put forth over the years (see section on the history of aging research for a timeline of central ideas from research). One focus has been on sirtuin (SIRT) enzymes of which there are 7 types in humans. SIRT 1, 6 and 7 are primarily in the cell’s nucleus, SIRT 3, 4, and 5 are in the cell’s mitochondria and SIRT2 is in the cell’s cytoplasm. Sirtuins are NAD+ dependent deacetylases. The sirtuin enzyme system is the start of a cascade of biological reactions that turn on and keep on cellular repair. Sirtuins are involved in the cell’s stress response system, DNA damage repair, maintaining genome integrity, positive regulation of telomere length, managing energy efficiency of metabolism and cause deacetylation of DNA by removing the + charge of DNA.
The change in DNA charge allows DNA to pack more tightly. More tightly packed DNA leads to reduced transcription (expression) of cellular DNA (as does other epigenetics such as methylation). This heterochromatin (packed DNA) is the more “youthful” chromatin state. This reversal to the heterochromatin state results in the stopping of inappropriate expression of normally silenced genes, non-coding transcriptional units, transposable elements (an example is Line 1 which has been linked to Diabetes type 1, Huntington’s disease and Schizophrenia), and endogenous viruses (example HIV-1) residing in DNA.
Initial observations, allowing focus
When we talk about changing the cell so that it “decides” to use energy for repair we are talking about “turning on” the enzymatic activity of sirtuin enzymes. That said, sirtuin enzymes turn on when they encounter and use NAD+ and cAMP. Both of these are molecules are cellular signals that the cell is out of energy. The observation that age reversal happens when the organism’s cells are out of energy was first seen by Clive McCay in 1935 and in 2000 it was observed by Lenny Guarente that this was the result of sirtuin enzymes and Shin Imai observed sirtuin enzymes used NAD+ as a co-substrate. In other words: no NAD+, no sirtuin activity. David Sinclair’s lab noted that: NAD+ regulates a pathway that coordinates nuclear-mitochondrial synchrony in response to changes in energy supply and oxygen levels. Lower NAD+ leads to lower communication between the cellular nucleus and the energy-producing mitochondria of cells which then leads to cell dysfunction and cellular aging.
The use of NMN instead of NAD+
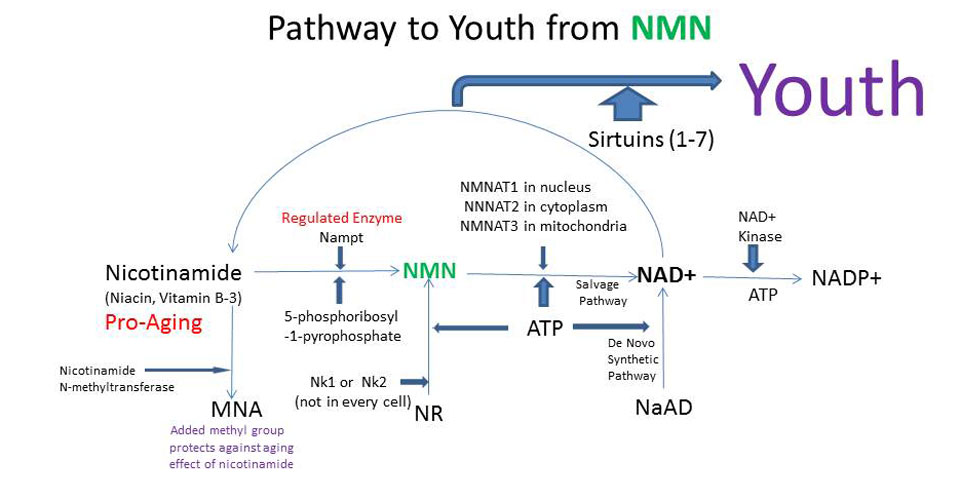
NAD+ is not absorbed through the gut, although can be taken up by IV drip, but even then only slowly. Nicotinamide mononucleotide (NMN) is dissolvable in water, is quickly absorbed by the gut, is a compound that naturally travels in the blood and is quickly turned into NAD+ by cells. NMN is normally made in the rate limiting the last step by salvage synthetic pathway that makes NAD+. Adding NMN increases NAD+ in cells because NMN is transformed with ATP (energy) by NMNAT (1,2,3) enzymes to make NAD+ in the nucleus (1), cytoplasm (2), and mitochondria (3) into NAD+.
Table: Mammalian Sirtuin Localization and Function
| Sirtuin | Localization | Activity | Examples of Targets |
|---|---|---|---|
| SIRT1 | Nucleus | Deacetylase | Telomeres, Histones, FoxO, PGC1α, p53, p300, C-Myc, NF-kB , PPAR |
| SIRT2 | Cytoplasm | Deacetylase | α-Tubulin, FoxO, PEPCK1, NF-kB |
| SIRT3 | Mitochondria | Deacetylase | ACS2, LCA, HMGCS2, GDH, IDH2, Ornithine transcarbamylase |
| SIRT4 | Mitochondria | ADP-ribosyltransferase | GDH |
| SIRT5 | Mitochondria | Demalonylase desuccinylase | CPS1 |
| SIRT6 | Nucleus | Weak deacetylase ADP-ribosyltransferase | Histones, PARP1, Hif1α, NF-kB |
| SIRT7 | Nucleus | Weak deacetylase | Histones, p53, Hif1α, Hif2α |